John Latimer filming in a SPRUCE experimental plot.
|
Fine-root and fungal dynamics in SPRUCE: 2012 - on-going
Manual minirhizotrons, along with
newly-developed automated minirhizotrons, were installed in each SPRUCE
experimental plot in fall, 2012. The manual minirhizotrons are being used to track the
responses of
fine-root dynamics to warming and elevated [CO2] over the next 10 years. The high-magnification and automated nature of the new minirhizotron systems from RhizoSystems, LLC, will allow us to track changes in root dynamics, as well as fungal hyphae, at much higher temporal resolution.
Collaborators: John Latimer, Joanne Childs
|

An array of ion-exchange resin-access tubes.
|
Plant-available nutrients in SPRUCE: 2013 - on-going
Several methods are
being used to track changes in plant-available NH4-N, NO3-N, and PO4 in response to warming and elevated [CO2] over the next 10 years.
We deployed WECSA ion-exchange resins across hummock-hollow
microtopography in two locations in each SPRUCE plot, and resins are
retrieved every 28 days during the growing season to determine an
integrated measure of root-available nutrients.
Collaborators: John Latimer, Joanne Childs, Deanne Brice, Holly Vander Stel
|

A root-ingrowth core with shrub roots and fungi.
|
Root ingrowth cores in SPRUCE:
2014 - on-going
Given the difficulty of distinguishing
between living and dead roots in the anaerobic bog environment, root
ingrowth cores constructed of rigid polypropylene tubes and filled with
commercial peat from a nearby bog are being
used to capture newly-grown roots. Ingrowth cores have been harvested
seasonally from paired hummock-hollow microtopography in two locations
in each SPRUCE plot. Cores
are sectioned into 10-cm increments, and spruce,
larch, and shrub roots are removed and analyzed by functional class for
mass, morphology, and chemistry (using jewelers glasses to see
shrub roots that are finer than a human hair!).
Collaborators: Deanne Brice, Joanne Childs, Holly Vander Stel
|

Ingrid and Kelsey on RPT (Root-Picking Thursdays).
|
Initial root, peat characteristics in SPRUCE:
2012
Initial root biomass distribution (along with peat characteristics and chemsitry) was determined
prior to initiation of experimental
treatments by taking 3-m deep soil cores in hummock-hollow
microtopography in each SPRUCE experimental plot in August, 2012.
Cores were sectioned into 10-cm or larger increments, and spruce,
larch, and shrub roots were removed (using jewelers glasses to see
shrub roots finer than a human hair!).
Iversen CM, Hanson PJ, Brice DJ,
Phillips JR, McFarlane KJ, Hobbie EA, Kolka RK. 2014. SPRUCE Peat Physical and Chemical Characteristics from Experimental
Plot Cores, 2012. Carbon Dioxide Information Analysis Center, Oak
Ridge National Laboratory, U.S. Department of Energy, OakRidge, TN, USA. http://dx.doi.org/10.3334/CDIAC/spruce.005.
|

Joanne filming roots in the S1 bog in 2011.
|
Fine-root dynamics in a bog:
2011 - 2012
Few experiments have investigated fine-root dynamics in wetland ecosystems. Prior to the SPRUCE experiment, we
installed minirhizotrons in an ombrotrophic bog ecosystem in northern MN,
USA across gradients of black spruce
density and hummock-hollow microtopography, and quantified root growth and mortality every week. Data are being
used to inform a future climatic and environmental change manipulation
on-site, and will add to our limited knowledge of rooting dynamics in
peatland ecosystems.
Iversen CM, Childs J, Norby RJ, Ontl TA, Kolka RK, Brice DJ, McFarlane K, Hanson PJ. Just beneath the surface: The distribution and dynamics of fine roots in an ombrotrophic bog. In preparation.
|
|

Minirhizotron images from the S1 bog over a period of
three weeks.
|
Advancing the use of minirhizotrons in wetlands
Together
with an international group of experts, we developed a consensus on,
and a methodological framework for, the appropriate installation and
use of minirhizotrons in wetlands.
Iversen CM, Murphy MT, Allen ME, Childs J, Eissenstat DM, Lilleskov EA, Sarjala TM, Sloan VL, Sullivan PF (2012). Advancing the use of minirhizotrons in wetlands. Plant and Soil 352: 23-39. [pdf]
See an ORNL press release on this paper, and associated news coverage at knoxnews.com and sciencedaily.com.
|
|
|
|
Next-generation Ecosystem Experiments (NGEE) - Arctic
*The goal of the NGEE - Arctic experiment (PI: Stan Wullschleger) is
to understand how the thawing of permafrost - and associated changes in
landscapes, hydrology, soil, and plants - affects feedbacks to the
climate
system. Phase I was focused on a gradient of polygonal tundra in
Barrow, AK (2011 - 2015), and in Phase 2 (2015 - 2018) we will build
upon work conducted in Phase 1 by establishing a more southerly site
characterized by transitional ecosystems, warm, discontinuous
permafrost, higherannual precipitation, and well-defined watersheds with strong topographic gradients.
... Phase 2 ...............................................................................................................................

Modeler, Peter Thornton, collecting information on Arctic
plant leaf traits.
|
Capturing the variation in plant traits across Arctic tundra: on-going research
One
of the five main tasks in Phase 2 will be to understand how warming and
permafrost thaw will affect above- and belowground plant functional
traits, and what the consequences might be for Arctic ecosystem carbon,
energy, water, and nutrient fluxes. This work will be conducted across
gradients of polygonal tundra in Barrow, AK, as well as in a more
southerly site near Nome, AK, on the Seward Peninsula.
Collaborators: A Breen, E Euskirchen, A Rogers, S Serbin, P Thornton, SD Wullschleger, C Xu , F Yuan
|
... Phase 1 ...............................................................................................................................

Injecting 15NH4Cl into the organic horizon in a patch
of Eriophorum angustifolium.
|
Differentiating arctic PFTs based on root nitrogen aquisition: on-going research
In July, 2013, we injected a 15N
tracer into the rooting zone at three targeted depth intervals (organic
layer, mineral soil, and the permafrost boundary) in order to characterize
nitrogen acquisition for species representing three important plant
functional types (PFTs) on
the Barrow Ecological Observatory. The amount of nitrogen acquired from
different depths throughout the soil profile is being used to refine
definitions of plant
functional types in arctic models.
Collaborators: J Childs, JA Liebig, RJ Norby, IJ Slette, VL Sloan, HM Vander Stel; SD Wullschleger
|

The lab team measuring soil characteristics prior to
the incubation.
|
Carbon and nutrient release from an organic-rich active layer across a gradient of polygonal tundra: on-going research
Using
soil cores collected in summer, 2012 in Barrow, AK, we simulated future
temperatures in a soil incubation study to investigate the effects of warming on soil carbon and nutrient mineralization.
Iversen CM,
Vander Stel HM, Norby RJ, Sloan VL, Childs J, Brice DJ, Keller JK, Jong
A, Ladd MP, Wullschleger SD. 2015. Active Layer Soil Carbon and
Nutrient Mineralization, Barrow, Alaska, 2012.Next Generation Ecosystem
Experiments Arctic Data Collection, Carbon Dioxide Information Analysis
Center, Oak Ridge National Laboratory, Oak Ridge, Tennessee, USA. Data
set accessed at http://dx.doi.org/10.5440/1185213.
Treat CC, Natali SM, Ernakovich J, Iversen CM, Lupascu M, McGuire AD, Norby RJ, Chowdhury TR, Richter A, Santruckova H, Schädel C, Schuur EAG, Sloan VL, Turetsy MR, Waldrop MP. 2015. A pan-Arctic synthesis of CH4 and CO2 production from anoxic soil incubations. Global Change Biology 21: 2787-2803. doi: 10.1111/gcb.12875.
|


Colleen (above) with the perfect soil core, Victoria (below)
clipping moss out of a clip plot.
|
Plant community dynamics across a gradient of polygonal tundra: on-going research
In
2012 and 2013, we
investigated the relationships among plant community composition,
above- and belowground biomass and N content, and edaphic and
environmental conditions, and how these relationships changed across a
gradient of
polygonal tundra in Barrow, AK.
Sloan VL, Brooks JD, Wood SJ, Liebig JA, Siegrist J, Iversen CM,
Norby RJ. 2014. Plant community composition and vegetation height,
Barrow, Alaska, Ver. 1. Next Generation Ecosystem Experiments Arctic
Data Collection, Carbon Dioxide Information Analysis Center, Oak Ridge
National Laboratory, Oak Ridge, Tennessee, USA. Data set accessed at http://dx.doi.org/10.5440/1129476.
Sloan VL, Liebig JA, Hahn MS, Curtis JB, Brooks JD, Rogers A, Iversen CM,
Norby RJ. 2014. Soil temperature, soil moisture and thaw depth, Barrow,
Alaska, Ver. 1. Next Generation Ecosystem Experiments Arctic Data
Collection, Carbon Dioxide Information Analysis Center, Oak Ridge
National Laboratory, Oak Ridge, Tennessee, USA. Data setaccessed at http://dx.doi.org/10.5440.1121134.
Sloan VL, Iversen CM, Liebig
JA, Curtis JB, Hahn MS, Siegrist J, Norby RJ. 2014. Plant Available
Nutrients, Barrow, Alaska Ver. 1. Next Generation Ecosystem Experiments
Arctic Data Collection, Carbon Dioxide Information Analysis Center, Oak
Ridge National Laboratory, Oak Ridge,Tennessee, USA. Data set accessed at http://dx.doi.org/10.5440/1120920.
|
|
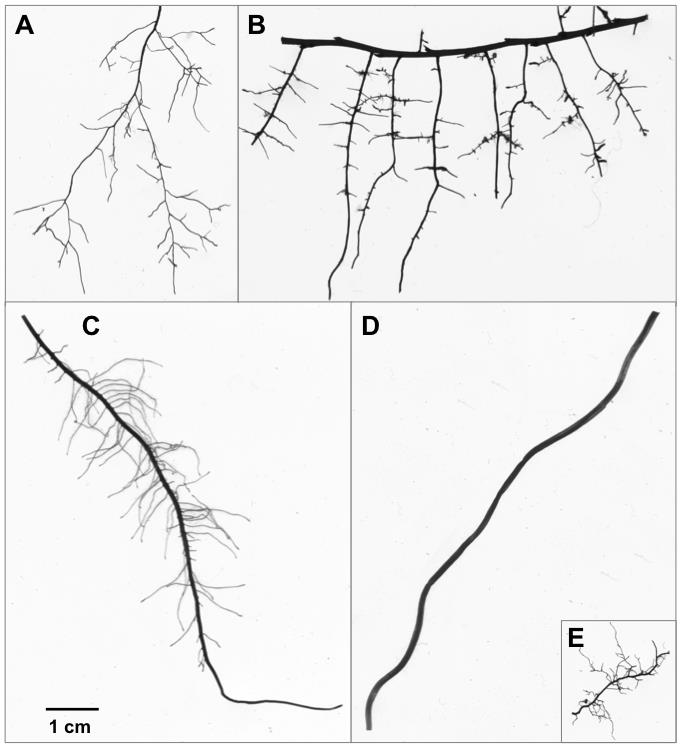
The unseen iceberg: Plant roots in arctic tundra
Plant
roots play a critical role in ecosystem function in arctic tundra, but
root dynamics in these ecosystems are poorly understood. To address
this knowledge gap, we synthesized available literature on tundra
roots, including their distribution, dynamics, and contribution to
ecosystem carbon and nutrient fluxes, and we highlighted key aspects of
their representation in terrestrial biosphere models.
Iversen CM,
Sloan VL, Sullivan PF, Euskirchen ES, McGuire AD, Norby RJ, Walker AP,
Warren JM, Wullschleger SD. 2015. The unseen iceberg: Plant roots in
arctic tundra (Tansley Review). New Phytologist 205: 34-58.
Iversen CM, Sloan
VL, Sullivan PF, Euskirchen ES, McGuire AD, Norby RJ, Walker AP, Warren
JM, Wullschleger SD. 2014. Plant Root Characteristics and Dynamics in
Arctic Tundra Ecosystems, 1960 - 2012. Next Generation Ecosystem
Experiments Arctic Data Collection, Carbon Dioxide Information Analysis
Center, Oak Ridge National Laboratory, Oak Ridge, Tennessee, USA. Data
set accessed at http://dx.doi.org/10.5440/1114222.
|
|
|
 Photo credit: J Brooks Photo credit: J Brooks
|
Belowground carbon partitioning in historical FACE experiment: on-going research
In summer 2011, we returned to the site of a historical Free-Air CO2-enrichment
(FACE) experiment in a sweetgum plantation. We girdled one-half of the
trees in each treatment plot with goals of determining the effects of
tree carbon inputs on soil carbon and nutrient cycling, and quantifying
the size of belowground carbon storage pools.
Collaborators: J Childs, CT Garten, RJ Norby, JM Warren
Lynch
DJ, Matamala R, Iversen CM, Norby
RJ, Gonzalez-Meler MA. 2013. Stored carbon partly fuels fine-root respiration
but is not used for production of new fine roots. New Phytologist 199:
420-430. [pdf]
See news coverage of our excellent student interns on a local TV station.
|

Photo credit: R. Norby
|
Carbon partitioning in a young loblolly pine
Our
objective was to improve the carbon partitioning routines in existing
ecosystem models, and test the models using short-term, comprehensive
field measurements of processes related to carbon partitioning from
leaves to roots and roots to soil.
Warren JM, Iversen CM, Garten CT, Norby RJ, Childs J, Brice DJ, Evans RM, Gu L, Thornton PE, Weston DJ (2012). Timing and magnitude of carbon partitioning through a young loblolly pine (Pinus taeda L.) stand using 13C labeling and shade treatments. Tree Physiology 32: 799-813. [pdf]
|
Oak Ridge National Laboratory Free-Air CO2-Enrichment experiment (ORNL FACE)
*A CO2-enriched sweetgum plantation located in Oak Ridge, TN, USA (1998-2009)

|
Belowground harvest: on-going research
We
excavated two large soil pits by hand in each treatment ring in late
June, 2009. Roots were separated into diameter classes for biomass and
nutrient analyses. Subsamples of sieved soil were used in a soil
incubation experiment to determine carbon and nitrogen mineralization
throughout the soil profile.
Iversen CM, Keller JK, Garten CT, Norby RJ (2012). Soil carbon and nitrogen cycling and storage throughout the soil profile in a sweetgum plantation after 11 years of CO2-enrichment. Global Change Biology 18: 1684-1697. [pdf]
Plant and soil samples have been archived and are available to the scientific community upon request. |

|
Root decomposition and carbon storage in soil organic matter: on-going research
We
combined root decomposition techniques with soil fractionation
techniques to quantify the transfer of carbon and nitrogen from
decomposing fine-root litter to relatively long-lived SOM using the
unique depleted C-13 signature of organic material in plants and soils
enriched with elevated [CO2]. We hypothesized that increased root quantity and decreased root quality under elevated [CO2]
would increase carbon and nitrogen storage in soil organic matter. This
research was funded by a dissertation improvement grant from the National Science Foundation.
Collaborators: JD Jastrow, RJ Norby |

|
Nitrogen cycling throughout the soil profile under elevated [CO2]
We
used isotope pool dilution to measure potential gross nitrogen cycling
rates throughout the soil profile. We found that nitrogen
mineralization at depth in the soil, combined with increased root
exploration of the soil volume under elevated [CO2], may be
more important than changes in potential gross nitrogen cycling rates
in sustaining forest responses to rising atmospheric CO2.
Iversen CM, Hooker TD, Classen AT, Norby RJ (2011). Net mineralization of N at deeper soil depths as a potential mechanism for sustained forest production under elevated [CO2]. Global Change Biology 17: 1130-1139. [pdf] |
|
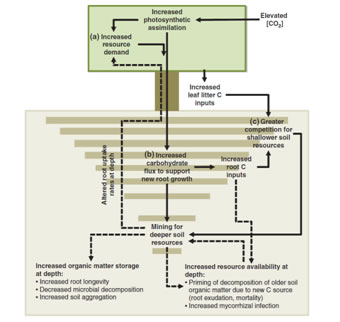
Digging deeper: Rooting distributions in CO2- enriched forests
Experimental evidence from a diverse set of forested ecosystems indicates that CO2-enrichment
may lead to deeper rooting distributions. Altered rooting distributions
are expected to affect important ecosystem processes such as root
physiology and soil nutrient cycling. However, the causes of greater
root production at deeper soil depths under elevated [CO2]
require further investigation. Progress in understanding and modeling
the interface between deeper rooting distributions and soil nutrient
cycling will be critical in projecting the sustainability of forest
responses to rising atmospheric [CO2].
Iversen CM (2010). Digging deeper: Fine root responses to rising atmospheric [CO2] in forested ecosystems. New Phytologist 186: 346-357. This paper was a finalist in the New Phytologist Tansley Medal competition. [pdf] |
|
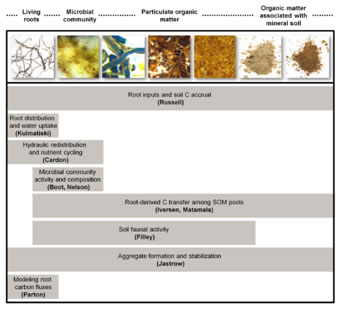
Missing links in the root-SOM continuum
Our goal was to synthesize root- and soil-centric studies into an
integrated understanding of belowground ecosystem processes in an
organized oral session at an annual ESA meeting. Speakers emphasized
the importance of the rhizosphere and soil environment for the
transformation of root-derived carbon to long-lived SOM. Integration of
observations made along the root-SOM continuum can lead to a more
holistic view of belowground ecology.
Iversen CM, O’Brien SL (2010). Organized Oral Session 3. Missing links in the root–soil organic matter continuum. Bulletin of the Ecological Society of America 91: 54-64. [pdf]
O ’Brien SL, Iversen CM (2009). Missing links in the root-soil organic matter continuum. New Phytologist 184: 513-516. [pdf] |

|
Root-derived carbon and nitrogen input to the soil
We
combined a long-term minirhizotron data set with continuous,
root-specific measurements to assess carbon and nitrogen input from
root mortality. We found that the flux of carbon and nitrogen into the
soil nearly doubled under elevated [CO2] due to stimulated
root production and mortality. Moreover, much of the carbon and
nitrogen input occurred relatively deep in the soil profile where
decomposition dynamics are likely to be different from what is commonly
observed and modeled in the upper soil.
Iversen CM, Ledford J, Norby RJ (2008). CO2 enrichment increases carbon and nitrogen input from fine roots in a deciduous forest. New Phytologist: 179: 837-847. [pdf] |

|
Nitrogen limitation controls carbon partitioning under elevated atmospheric [CO2]
In
a nitrogen fertilization experiment in an adjacent sweetgum stand, we
found that the increased production of ephemeral roots under elevated
[CO2] was most likely a mechanism for greater nitrogen
acquisition in response to nitrogen limitation within the stand. In
turn, the increased production of fine roots with a relatively large
nitrogen concentration necessitated increased nitrogen uptake from the
soil.
Iversen CM, Norby RJ (2008). Nitrogen limitation in a sweetgum plantation: Implications for carbon allocation and storage. Canadian Journal of Forest Research 38: 1021-1032. [pdf] |
Old-field Community Climate and Atmospheric Manipulation (OCCAM)
*Constructed old-field communities exposed to elevated [CO2], warming, and altered water conditions in open-top chambers located in Oak Ridge, TN, USA (2003-2008)

Photo credit: J. Childs
|
Fine-root dynamics in a constructed old-field
We
combined a long-term minirhizotron data set with continuous,
root-specific measurements to assess root biomass production and
mortality in a constructed old-field ecosystem exposed to elevated [CO2],
warming, and drought conditions. We asked whether multiple atmospheric
and climatic factors would interact to influence root production and
turnover throughout the soil profile.
Collaborators: J Childs, AT Classen, RJ Norby |
Nutrient-limited peatland ecosystems
*A fertilization experiment across an number of nutrient-limited bogs and fens located at the University of Notre Dame Environmental Research Center in the Upper Peninsula of Michigan, USA (1998-2002)

Photo credit: S. Bridgham
|
Scaling plant nutrient use in peatland ecosystems
We
fertilized a natural gradient of nutrient-limited peatland ecosystems
in Michigan, USA, with nitrogen, phosphorus, or a combination of both
nutrients. Our objectives were to determine how changes in carbon and
nitrogen partitioning within a plant, and changes in community
composition, would affect plant nitrogen-use efficiency. Plant
nitrogen-use efficiency and its components differed from the leaf- to
community level, and depended on nitrogen or phosphorus limitation.
Iversen CM, Bridgham SD, Kellogg LE (2010). Scaling plant nitrogen-use and uptake efficiencies in response to nutrient addition in peatlands. Ecology 91: 693-707. [pdf] |
|